导语
栽培烟草(Nicotiana tabacum)和本氏烟草(N. benthamiana)是植物生物学研究中广泛使用的模型,应用于植物与病原菌互作、代谢工程、功能基因组、合成生物学和基因编辑等研究中。栽培烟草 (2n=24) 和本氏烟草 (2n=19) 的多倍体基因组来自二倍体祖先的杂交。近十年前,有学者利用下一代测序技术(NGS)开发了栽培烟草N.tabacum的基因组草图(4.7 G),后有科学家基于光学图谱技术被改进到染色体水平的基因组,使其基因组N50达到了2.17 Mb,但染色体锚定率仅为64%。本氏烟草(N. benthamiana)的基因组草图有两个版本,大小分别为2.63 G和2.44 G。然而,由于缺乏高质量的参考基因组和注释,限制了这两种植物在植物和生物技术方面的巨大潜力。近日,《Molecular Plant》在线发表了一篇题为“High-quality assembled and annotated genomes of Nicotiana tabacum and Nicotianabenthamiana reveal chromosome evolution and changes in defense arsenals”的最新烟草基因组研究成果。让我们一起来学习一下吧~

文献标题:High-quality assembled and annotated genomes of Nicotiana tabacum and Nicotiana
benthamianareveal chromosome evolution and changes in defense arsenals
发表期刊:Molecular Plant
影响因子:27.5
发表时间:2024.01.24
研究思路
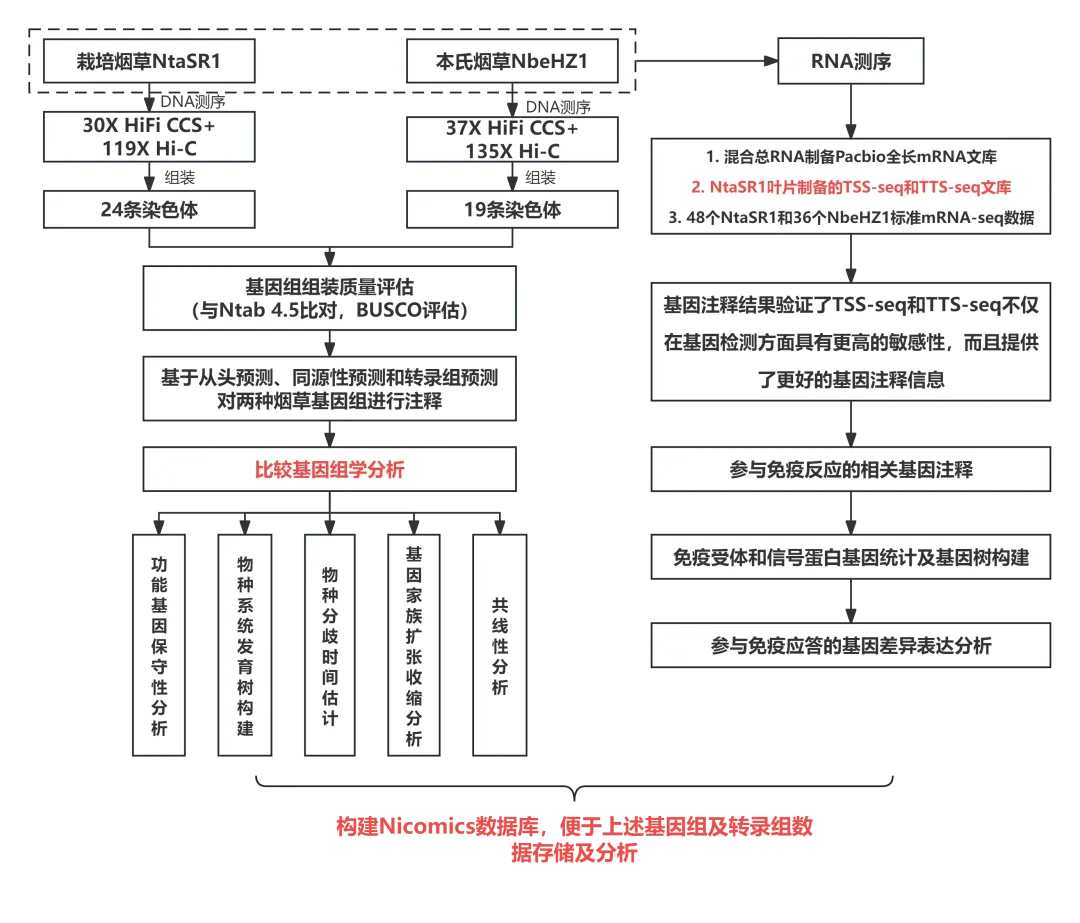
研究结果
1、本氏烟草和栽培烟草的高质量染色体水平完整基因组发现更多保守的功能基因
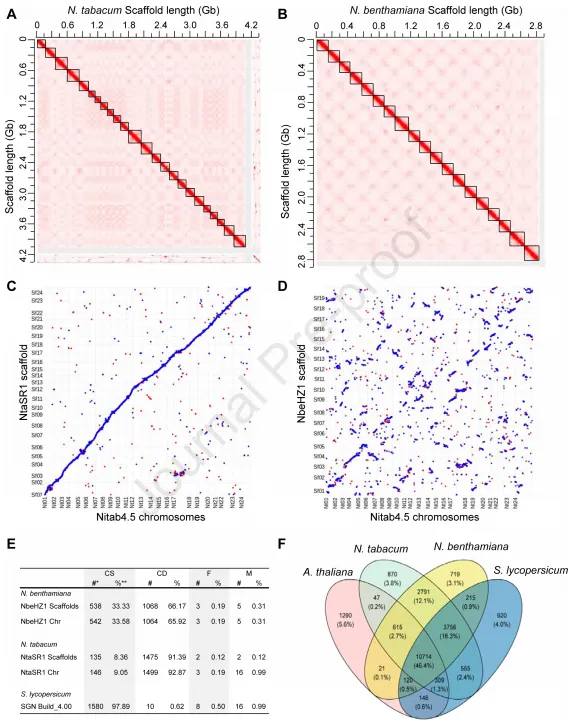
为了获得高质量的染色体水平的栽培烟草N. tabacum(Nta) 和本氏烟草N. benthamiana(Nbe) 参考基因组,对N. tabacum cv.Petite Havana SR1(NtaSR1)和来自华中农业大学的N. benthamiana实验室菌株(NbeHZ1)进行PacBio HiFi测序。总共获得了123 G (30X) 和105 G (37X) 的HiFi环状一致序列(CCS)读数,超过了在四倍体马铃薯中实现高质量单倍体特异性染色体组装的25X覆盖度。使用 hifiasm 软件组装了contig水平的基因组草图,Nta (4.3 G) N50 值为 56.07 Mb, Nbe (2.9 G) N50 值为 51.45 Mb。利用约 491 Gb Nta(119x)和 378 Gb Nbe(135x)的Hi-C 测序数据,将contigs锚定到染色体上,在对 Hi-C 数据进行过滤和映射后,92.65% 的 Nta 和 95.49% 的 Nbe contigs分别被锚定到 24 条和 19 条假染色体上。
为了评估组装基因组的质量,将本文组装的结果与N. tabacum 基因组 (Nitab 4.5) 进行了比对。NtaSR1 染色体与 Nitab4.5 染色体的共线性很强,组装率明显更高,然而,NbeHZ1 染色体与 Nitab4.5 染色体的共线性较弱。使用BUSCO进一步评估NtaSR1 和 NbeHZ1 基因组的完整性,分别检测到了1610 个(99.75%)和 1606 个(99.50%)完整的单拷贝(CS)或重复(CD)基因。这种高比率超过了组装良好的番茄基因组。这些结果表明组装的 NtaSR1 和 NbeHZ1 基因组质量很高。利用注释好的基因组,使用OrthoFinder对双子叶植物之间的功能基因进行了保守性分析,结果显示,拟南芥、番茄、NtaSR1 和 NbeHZ1 基因组中存在 10,714 个基因簇,占所有植物基因家族的 46.4%,与之前的基因组版本相比,发现了更多保守的功能注释基因,表明基因注释的完整性有所提高。
2、高质量的基因组序列揭示了N. tabacum和N. benthamiana的祖先以及这些物种的进化时间
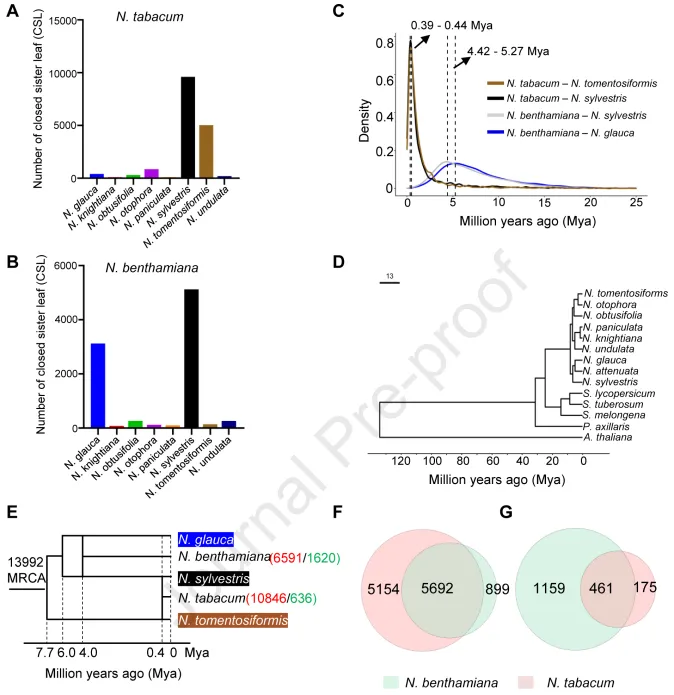
据报道,N. tabacum和N. benthamiana是由不同烟草属Nicotiana祖先物种杂交进化而来的四倍体。为了研究它们的基因组起源,对10个不同的烟草属物种进行注释和BUSCO分析,结果显示大多数烟草属物种的完整基因是单拷贝的,而N. rustica的基因是复制的,表明N. rustica是多倍体,而其他烟草属物种是二倍体。接下来,将NtaSR1 和 NbeHZ1 基因组与来自九个二倍体烟草属物种的同源基因池进行了比较。结果显示,NtaSR1 和 NbeHZ1 基因组分别与上述基因池共享 19,700 和 18,942 个基因。此外,利用 RAxML在 NtaSR1 和 NbeHZ1 同源物以及来自二倍体烟草属物种的同源物之间建立了闭合姊妹叶(CSL)系统发育树。该分析表明,NtaSR1 同源物与N. sylvestris和N. tomentosiformis同源物分别形成了约10,000 和 5,000 个 CSL。此外,在 NbeHZ1 同源物与N. glauca和N. sylvestris同源物之间分别观察到约 3,200 和 5,300 个 CSL。在这两种情况下,与潜在祖先形成的 CSL 数量是与非祖先形成的 CSL 数量的 6 至 100 倍。这些结果证明,N. tabacum和N. benthamiana分别起源于N. sylvestris和N. tomentosiformis的祖先以及N. glauca和N. sylvestris的祖先之间的杂交。
为了确定 N. tabacum 和 N. benthamiana 基因组进化的时间,计算了二倍体烟草属物种与 NtaSR1 或 NbeHZ1同源基因之间的同义替换率(Ks)。Ks 密度分布的分析表明,NtaSR1 基因组出现于 0.39 至 0.44 百万年前(Mya),而 NbeHZ1 基因组出现于 4.42 至 5.27 Mya。此外,利用二倍体烟草、番茄、马铃薯、茄子、矮牵牛和拟南芥基因组构建了系统发生树。结果表明,茄属植物和烟草之间的分化发生在 23 Mya 左右,而N. sylvestris和N. tomentosiformis以及N. sylvestris和N. glauca之间的分化分别发生在 7 Mya 和 6 Mya 左右。因此,该结果表明,N. benthamiana 基因组在两个亲本物种分离后很早就出现了,而 N. tabacum 基因组在亲本物种分化后的较晚时期才出现。分析还显示,在烟草属祖先物种中有 13,992 个最新共同祖先(MRCA)基因家族。N.benthamiana多倍体化后分别有 6,591 和 1,620 个基因家族发生了基因扩增和基因收缩,N.tabacum多倍体化后分别有 10,846 和 636 个基因家族发生了基因扩增和基因收缩。N. benthamiana基因组中扩大的基因家族在很大程度上与N. tabacum基因组中扩大的基因家族重叠,但远小于N. tabacum基因组中扩大的基因家族,而N. benthamiana基因组中收缩的基因家族数量大于N. tabacum基因组中收缩的基因家族数量。这些结果表明,基因扩增主要是由多倍体化引起的。N. benthamiana中观察到的基因扩张较少和基因收缩较多的现象与该物种在多倍体化后出现了严重的染色体丢失是一致的。
3、N. tabacum和N. benthamiana基因组染色体水平的结构变化和进化利用高质量的染色体水平的基因组组装结果
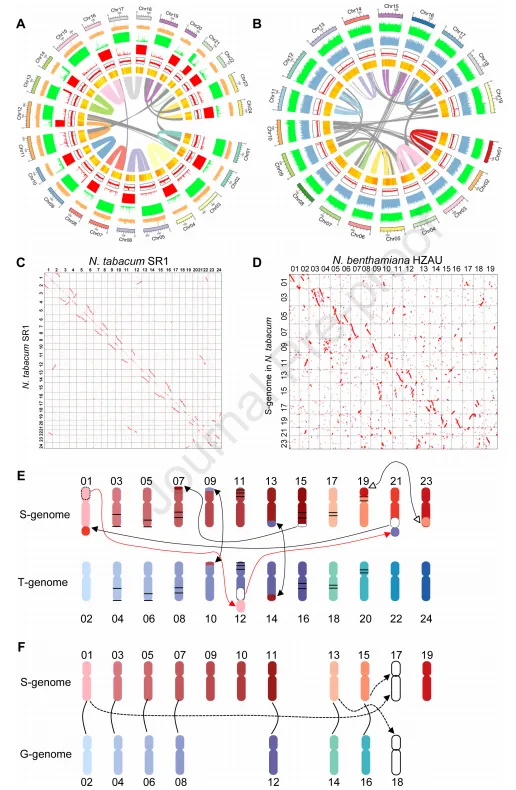
进一步研究了导致N. tabacum和N. benthamiana基因组形成的亚基因组之间和内部的染色体结构变化。NtaSR1和番茄染色体同源单拷贝基因比对结果表明,再茄属和茄科植物多样化以后,发生了大规模的染色体重排现象。将重新排序的NtaSR1与番茄染色体进行比对,明确了NtaSR1每条染色体的亲本起源,与N. tabacum基因组的最近的演化一致。并且,所有染色体上都发生了同源染色体之间的小片段交换,但S-(N. sylvestris)和T-(N. tomentosiformis)基因组之间的大片段染色体交换仅出现在部分染色体(9、10、12、13、14和21)上。此外,几对同源染色体中,3-4、5-6、7-8、15-16和17-18对中的一个染色体上存在大片段染色体倒位。S-基因组染色体的顶端片段与T-基因组染色体的底部片段呈相反方向比对,表明S-基因组染色体的底部被转位到了染色体7的顶部,然后发生了倒位。
NbeHZ1染色体与其祖先N. sylvestris和N. glauca的比对显示,两个亲本基因组在所有染色体上的分布均匀。将组装后的NbeHZ1基因组与有着共同祖先的NtaSR1 S 基因组进行了比对,当两条染色体与同一条NtaSR1染色体共线性最高时,被视为同源染色体。分析结果显示,NbeHZ1 染色体对 chr1-2、chr3-4、chr5-6、chr7-8、chr11-12 和 chr15-16 与 NtaSR1 染色体有明显的共线性,并有明显的亲本来源。同样,chr9 和 chr10 与 NtaSR1 chr09 和 chr11 有明显的共线性,但它们的同源染色体丢失了。NbeHZ1 chr15 和 chr21 的共线片段分布在其 chr2、9、10、11 和 12 中,表明这两对亲本染色体丢失了。
总之,该文章对改良后的基因组的详细分析揭示了当前N. tabacum和N. benthamiana基因组形成的大规模染色体水平结构变化的分子特性。
研究总结
该研究报告了本氏烟草和栽培烟草染色体水平参考基因组的构建情况,估计其完整性分别为99.5%和99.8%。开发了敏感的转录起始和终止位点测序方法,并将其用于栽培烟草的精确基因注释。比较基因组学分析揭示了导致各物种杂交基因组形成的亲本起源和染色体结构变化的证据。有趣的是,在本氏烟草中,抗病毒沉默基因 RDR1、RDR6、DCL2、DCL3 和 AGO2 从一个或两个亚基因组中消失,而在栽培烟草中则保留了两个同源基因。此外,与栽培烟草相比,本氏烟草基因组编码的免疫受体和信号转导元件较少。这些发现揭示了本氏烟草超易感特性背后的原因。并且,该研究开发了用户友好的 Nicomics (http://lifenglab.hzau.edu.cn/Nicomics/)网络服务器,便于基因结构和表达分析。
参考文献
Wang J, Zhang Q, Tung J, Zhang X, Liu D, Deng Y, Tian Z, Chen H, Wang T, Yin W, Li B, Lai Z, Dinesh-Kumar SP, Baker B, Li F. High-quality assembled and annotated genomes of Nicotiana tabacum and Nicotiana benthamiana reveal chromosome evolution and changes in defense arsenals. Mol Plant. 2024 Jan 24:S1674-2052(24)00008-X. doi: 10.1016/j.molp.2024.01.008. Epub ahead of print. PMID: 38273657.
