耶鲁大学的阿兰·韦纳和南希·梅泽尔斯夫妇综合了许多拼图碎片般的分子生物学事实,提炼出了一个基因组标签假说。这里将要介绍其中最关键的几块碎片,它们不只与假说有关,也是展现生命活动精密调节的好例子。
这些碎片中的第一块藏在芜菁黄花叶病毒(Turnipyellowmosaicvirus,TYMV)里。顾名思义,这种病毒会感染芜菁,也就是俗称蔓菁、盘菜、大头菜的那种根用蔬菜,使它的叶片出现黄色的斑。
除此之外,它也会感染其他的芸薹属蔬菜,比如卷心菜、大白菜、小白菜、花椰菜。芜菁黄花叶病毒发现于20世纪50年代,是一种非常典型的RNA病毒。它的整个基因组就是一条长约6300个碱基的单链RNA通过自我配对,这条RNA在三维空间中形成了非常复杂的结构。
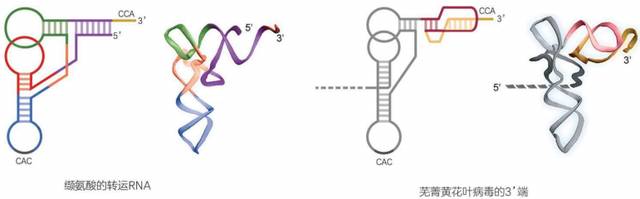
不过我们并不需要观察它的全部形态,只集中观察它的3’端末尾,就足够惊喜了。看到了吗?这个病毒的3’端结构,竟然与转运RNA惊人地类似!它不但有3’端的CCA尾,甚至还有缬氨酸的反密码子“CAC”。

只是转运RNA的迷你螺旋由5’端和3’端互相缠绕而成,但芜菁黄花叶病毒的5’端延抻出去编码基因了,所以只能靠3’端单枪匹地制造这个迷你螺旋。于是,这个3’端像小麻花似的扭了一圈,硬是凑出了酷似迷你螺旋的结构。
这个结构乍看很像打结,却又没有彼此穿过,不能真的系紧,所以在RNA的形态研究中被叫作“假结”。事实证明,这个“转运RNA样结构”和真的转运RNA一样好用。芜菁黄花叶病毒一旦在细胞内大量复制,缬氨酸的aaRS就会把病毒的转运RNA样结构当作真的转运RNA,在它们的CCA尾上连接一个缬氨酸,结果真正的转运RNA就得不到足够的缬氨酸,正常的蛋白质翻译工作因此缺乏原料,于是,叶片就发黄了,斑斑驳驳的。
可是,芜菁黄花叶病毒为什么要在3’端长出这样一个转运RNA样结构,和细胞争夺缬氨酸呢?因为只有给假结带上这个缬氨酸,芜菁黄花叶病毒才能迅速地复制,扩大感染,否则,如果那个转运RNA样结构发生了什么突变,抢不到足够的缬氨酸,病毒的扩增速度就会大幅减缓,甚至无法形成感染。
不过另一方面,人工编辑那个转运RNA样结构,把它们改成结合甲硫氨酸,新的芜菁黄花叶病毒也仍然能够感染大白菜,所以它们有可能只是必须结合一种氨基酸,而与具体是哪种氨基酸关系不大。
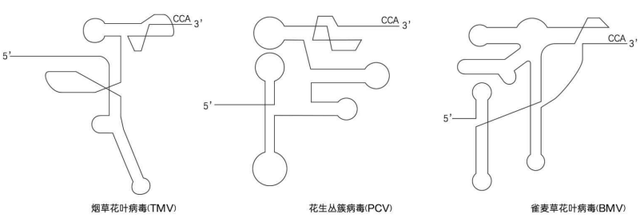
芜菁黄花叶病毒当然不是孤例,从20世纪70年代开始,人们陆续发现,一大批专门感染植物的RNA病毒都有这个转运RNA样结构,虽然未必都有完整的反密码子,但通过各种假结的堆叠,形状上都与真正的转运RNA非常类似,都能骗得细胞把氨基酸交给它们——除了缬氨酸,苏氨酸和组氨酸也是RNA病毒努力争夺的氨基酸。
而且,这些病毒的转运RNA样结构也往往与它们的复制能力有关。比如烟草花叶病毒,就是这一幕引文里的那种病毒,就必须要有这个结构才能顺利地结合RNA复制酶,如果把烟草花叶病毒的转运RNA样结构安装到一般的信使RNA上,这个信使RNA的表达效果竟然会增强100倍!
不仅如此,细胞内部有很多“RNA外切酶”,它们会从两端开始,将所有来源不明的RNA粉碎掉,而这个转运RNA样结构同样是破解之道。
转运RNA样结构只要连接上氨基酸,另外一类被称为“延伸因子”的蛋白质就会结合上去,包住病毒仿造的迷你螺旋,它们的本职工作是协助完全成熟的转运RNA进入核糖体,加快翻译的速度,但在此时,它们成了病毒3’端的保护套,可以阻止细胞内的RNA外切酶攻击这个病毒。
看到这里,读者或许会问:基因组标签假说谈论的是半个转运RNA可以引导复制,但这些病毒的转运RNA样结构却酷似一个完整的转运RNA,这岂不是有些矛盾吗?
这就引出了一个有趣的事实:早在20世纪90年代,我们就发现这些转运RNA样结构只需要假结那一半就能从aaRS那里骗来氨基酸,这与转运RNA只需迷你螺旋那一半就能与aaRS作用是一模一样的。
所以,如果说转运RNA样结构与转运RNA一样古老,经历过一样的进化,这并不是什么不可能的事情。毕竟我们早就说过,RNA病毒是非常古老的病毒,保不齐就是RNA世界的孑遗。但是,这“一样的进化”究竟是怎样的进化呢?
直观地看,一种可能是“拟态”,就像眉兰的花朵进化得酷似雌蜂,可以诱骗雄峰前来传粉那样,病毒的3’端本来与转运RNA没有任何关系,但是3’端长得越像转运RNA,就越能诱骗aaRS,捞到额外的复制优势。
所以早在迷你螺旋刚刚出现的时候,这些病毒的3’端就进化成了假结来占便宜,后来迷你螺旋进化成了转运RNA,这些假结也亦步亦趋,跟着进化成了转运RNA样结构。相对的,另一种可能是“平行”,就像牛和马的祖先都有蹄子,它们一直都在同样的草原上奔跑,所以也一直都有类似的蹄子。
或许,当初的RNA世界本来就既有迷你螺旋又有假结,它们从那时起就在共享原始的aaRS。后来,迷你螺旋被细胞征用,进化成了转运RNA,假结却成了病毒的工具,进化成了转运RNA样结构,但它们一直都在利用aaRS,所以一直保持着形态上的高度一致。
如果只看这种现象本身,那么“拟态”的确是最合理的解释,因为拟态作为极致的伪装,的确是生存竞争中极其常用的伎俩,我们可以在今天的世界里找到数不清的案例。但我们又不能只看这种现象本身,还有其他密切联系的碎片同样正摆在我们面前。第二块碎片,藏在发霉的面包里。
粉红面包霉菌(Neurosporacrassa)是1843年在法国巴黎的一家面包店里发现的,被它们感染的面包会迅速蔓延起蓬松、粉嫩、好像猪肉松一样的菌丝,对于19世纪的欧洲人来说,这样的面包反正是不能吃了,得赶紧丢掉,免得污染了那些新鲜的面包。但是到了20世纪,这些霉菌却成了实验室里的宝贝。

因为它们生长迅速,而且菌丝都是单倍体,没有显性基因遮盖隐性基因的麻烦,正是研究基因与性状的良好模型,所以直到今天,它们都是遗传学和细胞生物学上相当重要的模式生物。
不过,这种霉菌与基因组标签假说的联系和它们的单倍体特征一点儿关系都没有,因为是不是单倍体是细胞核里的事情,而我们关心的是这种霉菌的线粒体里的逆转录质粒。
一般来说,质粒是一些环状的小型双链DNA,自主地复制出许许多多份,分散在细胞的基质中。逆转录质粒也是如此,唯一不同的是,其他质粒都是通过DNA聚合酶直接复制自己,逆转录质粒却是先转录成RNA,再逆转录回DNA的。
而这些质粒最令韦纳夫妇感到兴奋的是,它转录出来的RNA的3’端竟然也是一个带着CCA尾的迷你螺旋,它会展现出酶RNA的本领,在CCA尾的末端自我剪切,使每一个单体互相分开。

不仅如此,这些RNA单体还可以发挥信使RNA的功能,在核糖体上翻译出一些蛋白质来,这其中又包括一个专门的逆转录酶,这个逆转录酶会专门识别这个迷你螺旋,结合上去,从CCA开始把整条RNA逆转录成许许多多环状的双链DNA,也就是许许多多新的逆转录质粒。也就是说,逆转录质粒的RNA可以利用自身的迷你螺旋启动逆转录。
如果你觉得这与转运RNA的迷你螺旋联系不够直接,还只是“像”而已,那么不妨往下看,在进一步的实验中,人们发现这两种质粒的逆转录酶真的可以结合转运RNA的迷你螺旋,然后从那里开始,把整个转运RNA都逆转录成DNA。XXIX既然已经谈到了“逆转录”,韦纳夫妇的第三块碎片的确藏在逆转录病毒里。
几乎所有的逆转录病毒,都要劫持宿主的转运RNA,用那个迷你螺旋上的CCA尾启动逆转录。最大名鼎鼎的逆转录病毒当然就是HIV,这是一种RNA逆转录病毒,基因组记录在一条单链RNA上。
当它们嵌入人体的细胞,就要利用自身携带的逆转录酶,把这条单链RNA逆转录成一条双链DNA,插进细胞自身的染色体中,从此再也无法被免疫系统清除。其中,把单链RNA逆转录成双链DNA是一个非常复杂的过程。

在这里,我们只需记住一件事:HIV的逆转录酶不能凭空逆转录一条RNA,而必须有个什么东西事先结合在RNA上,给它开个头,而那个“东西”,就是转运RNA的迷你螺旋,它们开的头,就是那个CCA。
另外,逆转录病毒也不都是以RNA为遗传物质,比如乙肝病毒和花椰菜花叶病毒(CaMV),它们都是逆转录病毒,但遗传物质却是环状的双链DNA。进入宿主的细胞后,它们会先把自己的DNA转录成单链的RNA,再把单链的RNA逆转录成环状的双链DNA,整个过程与逆转录质粒的复制过程非常类似。
但不同的是,它们转录出来的RNA自己没有迷你螺旋,必须像HIV一样盗用宿主的转运RNA,启动逆转录。第四块关键的拼图碎片,就藏在我们自己的身体内,它就是端粒酶。
人类以及真核域的一切成员最主要的遗传物质都是线状的双链DNA,它们被封装在一条条的染色体里,通过一次次的细胞分裂世代传递。但是,细胞的DNA复制系统有一种先天缺陷:DNA的头尾两端都会有一小截复制不下来,直接丢失掉,复制的次数多了,整个DNA就散架了。